大家好,这里是专注表观组学十余年,领跑多组学科研服务的易基因。
近日,中国农业科学院蔬菜花卉研究所杨学勇研究员、中国农业科学院深圳农业基因组所黄三文研究员和英国约翰英纳斯中心丁一倞研究员团队合作,以封面文章形式在顶刊《Cell》(细胞)上发表题为“Recessive epistasis of a synonymous mutation confers cucumber domestication through epitranscriptomic regulation”的突破性前沿成果。研究聚焦黄瓜果实长度的驯化性状,发现两个紧密连锁且存在上位互作的基因——FL1.1和FL1.2,分别由 YTH1 和 ACS2 基因定义,揭示了ACS2中的1287位C>T 同义突变(不改变氨基酸序列的DNA变异)通过介导m6A修饰变化和mRNA结构构象调控ACS2蛋白翻译效率(TE),进而影响黄瓜果实长度的驯化机制,为理解同义突变的生物学功能提供了多细胞生物层面实例。

标题:Recessive epistasis of a synonymous mutation confers cucumber domestication through epitranscriptomic regulation(同义突变的隐性上位效应通过表观转录组调控赋予黄瓜驯化特性)
发表时间:2025年8月21日
发表期刊:Cell
影响因子:IF42.5/Q1
技术平台:遗传定位与基因克隆、基因编辑、m6A-seq、RNA结构分析、免疫印迹与核糖体图谱分析等
作者单位:杨学勇研究员、黄三文研究员和丁一倞研究员为本文共同通讯作者,中国农业科学院蔬菜花卉研究所辛同旭博士、河南大学张震副教授、英国约翰英纳斯中心张月莹博士和中国农业科学院蔬菜花卉研究所已毕业硕士生李旭彤为本文共同第一作者。
DOI:10.1016/j.cell.2025.06.007
长久以来,同义突变(Synonymous mutations)因不改变蛋白质功能而被视为“沉默突变”。近期研究发现同义突变可能影响转录或转录后过程,但其对生物性状的影响,尤其是在个体水平上的影响,仍有待深入研究。本研究鉴定出两个紧密连锁且存在上位互作的基因:YTH1(m6A reader蛋白编码基因)和ACS2(ACC合成酶编码基因),这两个基因与黄瓜果实长度驯化高度相关。ACS2中的致病突变是1287位C>T同义突变。具体而言,在野生黄瓜(hardwickii)中,ACS2¹²⁸⁷C会导致腺苷残基发生m6A修饰,并导致松散的RNA结构构象,从而提高ACS2蛋白表达水平,使果实变短;在栽培黄瓜(Xintaimici,新泰密刺)中,ACS2¹²⁸⁷T 会破坏m6A甲基化,形成紧密的RNA结构构象,导致蛋白表达量降低,果实变长。本研究为表观转录组调控通过同义变异影响生物性状提供了遗传学证据。
研究亮点
- 两个存在上位互作的基因YTH1和ACS2赋予黄瓜驯化特性。
- ACS2的1287位C>T同义突变是致病变异。
- 1287T破坏邻近的m6A甲基化,并形成紧凑的ACS2 RNA结构。
- m6A使RNA结构构象更弱,并增强翻译效率。
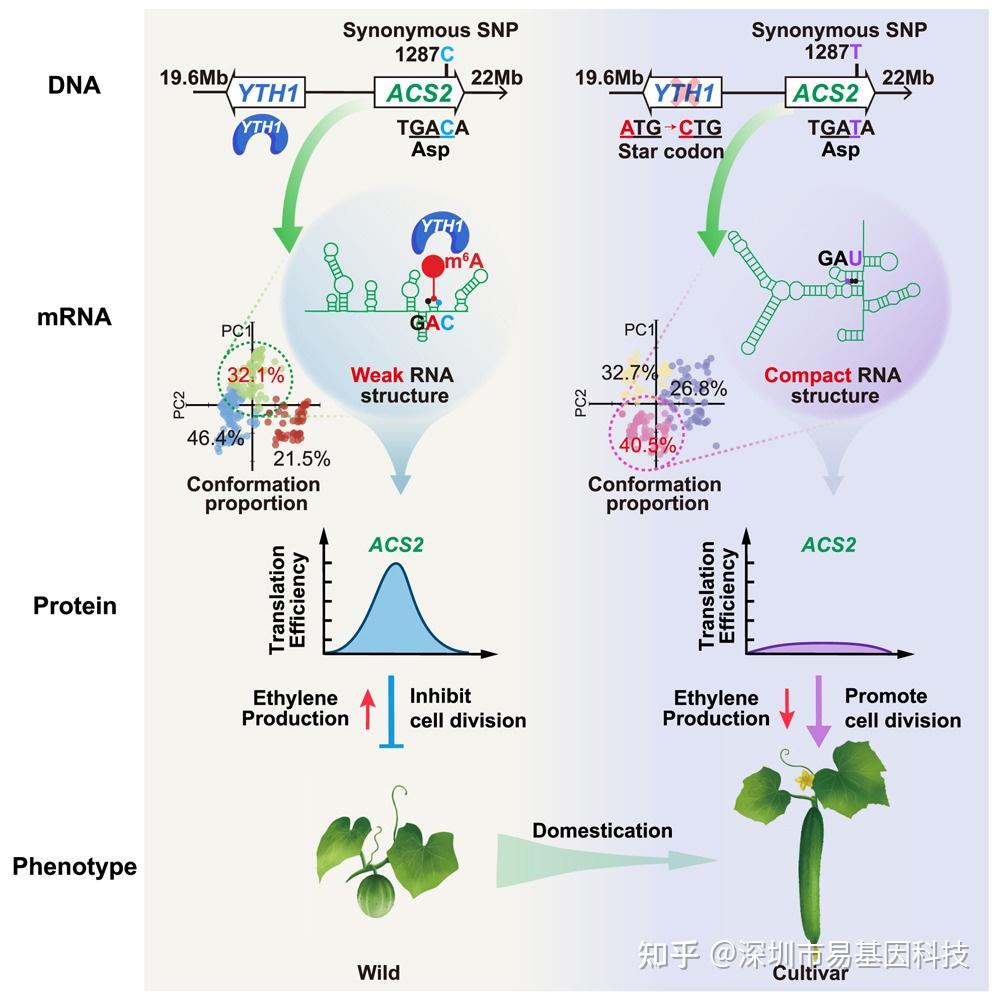
研究图形摘要
易小结
本研究通过 m6A-seq 等技术揭示了黄瓜果实长度驯化的遗传机制,证明了ACS2基因中的1287位C>T同义突变通过改变m6A修饰和mRNA结构调控ACS2蛋白的翻译效率,进而影响果实长度。
m6A-seq在本研究中不仅揭示了ACS2 mRNA上的m6A修饰位点及其在驯化过程中的变化,还进一步阐明了m6A修饰与RNA结构、翻译效率之间的复杂关系。这不仅为作物驯化研究提供了新的思路,也为易基因的表观基因组学技术在农业生物领域的应用提供了重要参考。易基因的表观基因组技术,如m6A-seq、ChIP-seq和WGBS等,能够全面解析基因表达调控网络,助力研究人员深入理解生物性状的遗传基础,为作物改良和新品种培育提供科学依据。
易基因相关产品拓展性案例展示
- 植物抗逆:MeRIP-seq等揭示mRNA m6A甲基化调控拟南芥的抗寒性分子机制
- 作物育种:MdMTA介导的RNA甲基化(m6A修饰)在苹果抗逆品种选育中的作用研究
- 项目文章|IJBM:安医大陈飞虎团队揭示METTL3介导m6A甲基化在炎症性疾病发病机制中的表观调控作用
- 项目文章|EP/IF7.3:陆军军医大学刘晋祎团队揭示PM2.5暴露通过m6A甲基化调控雄性生殖功能损伤机制
- 项目文章 | 贵州省医刘代顺团队MeRIP-seq揭示m6A修饰在病毒感染中的免疫调控作用
- 项目文章 | MeRIP-seq揭示m6A修饰在肺动脉高压(PAH)发病机制中的潜在作用和新治疗靶点
研究方法
- 遗传定位与基因克隆:利用野生黄瓜(hardwickii)和栽培黄瓜(Xintaimici)构建回交导入系(introgression lines,ILs),通过精细定位将调控果实长度的FL1位点细分为FL1.1和FL1.2两个紧密连锁的基因位点,并进一步克隆出这两个位点的关键基因YTH1和ACS2。
- 基因编辑:开发适用于瓜类作物的单碱基编辑工具,将ACS2野生等位基因的1287位的C编辑为T,直接验证该同义突变的遗传功能,证明其对黄瓜果实长度驯化的调控作用。
- m6A-seq和m6A-RIP-qPCR:通过m6A-seq分析ACS2 mRNA的m6A修饰水平,发现ACS2 mRNA的第三外显子存在显著的m6A peaks,且1287位C>T同义突变显著降低ACS2 mRNA的m6A甲基化水平,揭示m6A修饰在调控ACS2蛋白翻译效率中的关键作用。
- RNA结构分析:采用单分子RNA结构构象多样性分析(DAVINCI)等技术,分析ACS2 mRNA的结构构象多样性,发现1287位C>T同义突变介导mRNA结构从松散状态变为紧凑状态,进而影响YTH1与ACS2 mRNA的结合能力及ACS2翻译效率。
- 免疫印迹与核糖体图谱分析:通过免疫印迹检测不同基因型植株中ACS2蛋白的水平,结合核糖体图谱分析ACS2的翻译效率(TE),验证1287位C>T同义突变及YTH1对ACS2蛋白水平和TE的调控效应。
结果图形
(1)与果实长度驯化相关的两个紧密连锁的互作基因
研究人员发现黄瓜果实长度这一驯化性状由位于1号染色体的FL1位点调控,进一步将该位点细分为FL1.1和FL1.2两个紧密连锁的基因位点。通过构建回交导入系(ILs)和F₂群体分析,发现FL1.1和FL1.2基因位点存在上位互作,且FL1.2对FL1.1位点具有隐性上位性效应。当FL1.2为栽培等位基因时,无论FL1.1的基因型,果实始终是长瓜。这表明黄瓜果实长度的驯化由这两个紧密连锁且互作的数量性状位点(QTL)共同决定。
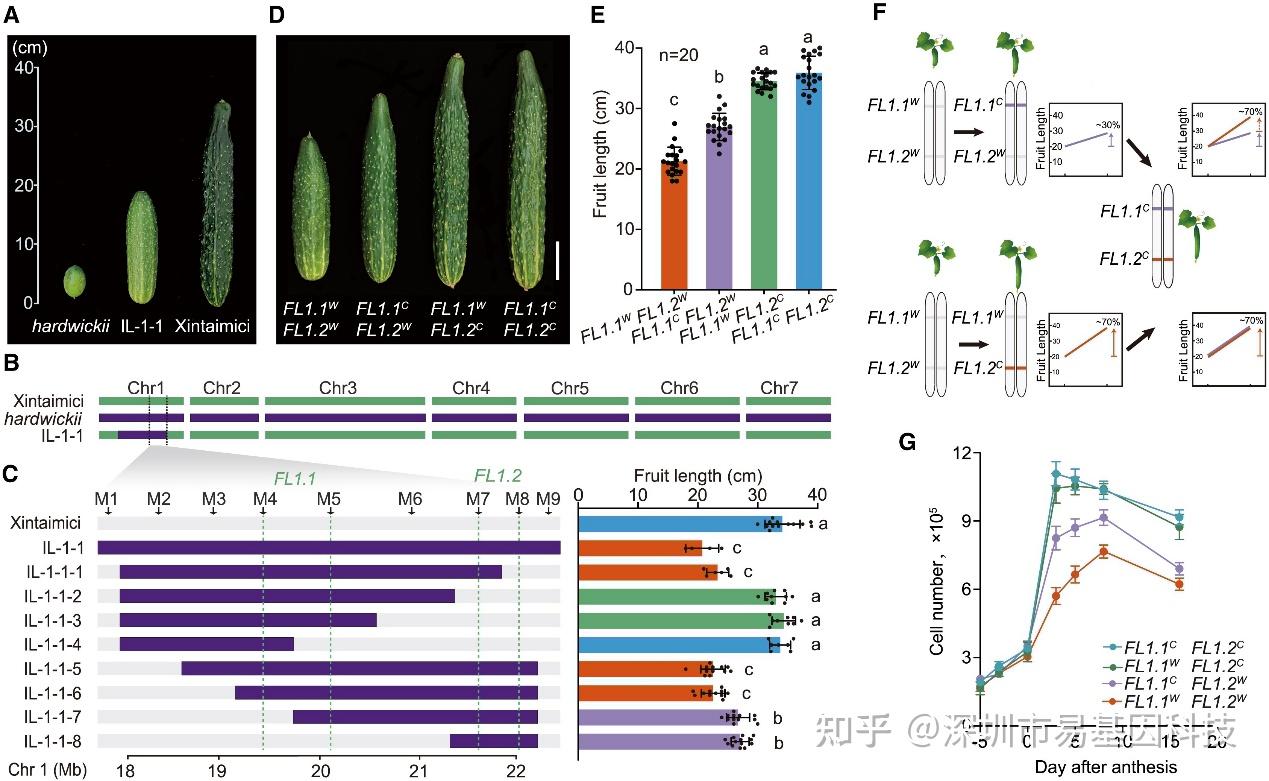
图1:与果实长度驯化相关的两个紧密连锁、相互作用的基因
(A)野生黄瓜hardwickii(左侧)、栽培黄瓜Xintaimici(右侧)以及IL-1-1(中间)的代表性果实。
(B)代表性ILs和亲本基因型。绿色和紫色分别代表Xintaimici和hardwickii的基因型。
(C)通过将IL-1-1渐渗片段细分为更小的片段(IL-1-1-1至IL-1-1-8),鉴定并定位两个互作位点FL1.1和FL1.2。左侧为选定重组体的基因型。灰色和紫色分别代表Xintaimici和hardwickii的基因型。绿色虚线定义了FL1.1和FL1.2的遗传区间。右侧为相应重组体植株的果实长度。
(D)四种基因型的代表性果实表型。W和C上标分别代表野生型和栽培型等位基因。
(E)与(D)相关的果实长度定量分析。
(F)模式图显示FL1.2C在遗传上是隐性的,并对FL1.1具有上位性效应。紫色和红色分别代表FL1.1C和FL1.2C的基因型。
(G)四种基因型的细胞数量分析。
(2)FL1.2由ACS2基因的同义突变定义
研究团队将FL1.2定位到17.3kb区间,发现该区间仅含ACS2基因。ACS2编码1-氨基环丙烷-1-羧酸合成酶,是乙烯合成的限速酶。基因组测序显示,野生黄瓜和栽培黄瓜的ACS2存在多个变异,但只有1287位C>T同义突变显著影响ACS2蛋白水平。通过双荧光素酶实验和转基因实验验证,1287位C>T同义突变降低了ACS2蛋白水平,进而导致黄瓜果实变长,是FL1.2调控黄瓜果实长度驯化的因果突变。
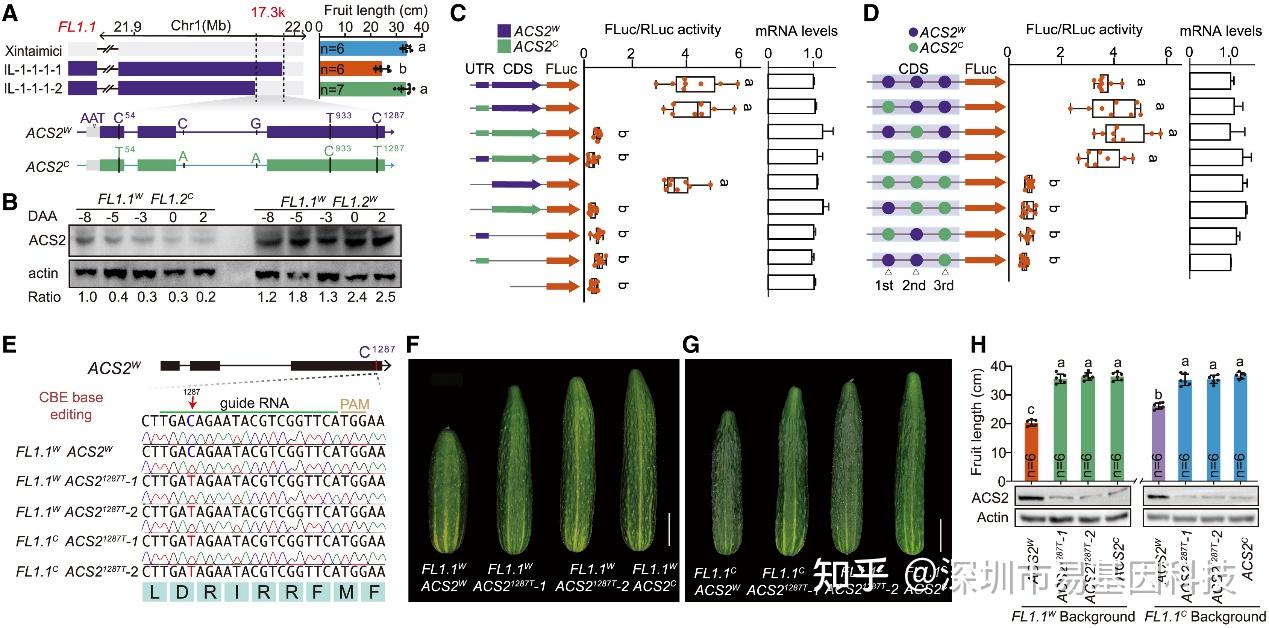
图2:FL1.2由ACS2基因的同义突变定义
- FL1.2的精细定位区间包含ACS2基因。上:选定重组体的基因型和果实长度。下:ACS2W和ACS2C的基因结构和变异。方框、线条和灰色方框分别代表外显子、内含子和非翻译区(UTRs)。
- 免疫印迹分析显示早期果实发育过程中ACS2蛋白的水平。
- 双荧光素酶报告基因分析对ACS2的5′UTR或CDS变异进行功能鉴定。左侧:示意图显示ACS2W和ACS2C的5′UTR和CDS变异的所有组合构建。中间:左侧显示相应构建的FLuc/RLuc活性。右侧:相应构建的相对ACS2 mRNA表达水平。
- 使用双荧光素酶报告基因分析法对三个同义SNP进行功能鉴定。左侧:示意图显示ACS2W和ACS2C中三个同义SNP的不同组合构建。中间:左侧显示相应构建的FLuc/RLuc活性。右侧:各种构建的相对ACS2 mRNA表达水平。
- ACS2¹²⁸⁷C的碱基编辑结果。
(F-G) 带有C1287到T转换的转基因植株果实长度。
(H) 与(F)和(G)相关的果实长度和ACS2蛋白水平的定量分析。
(3)FL1.1由YTH1基因的有害突变定义
FL1.1被定位到11kb区间,该区间仅含YTH1基因。YTH1编码m6A reader蛋白。基因组测序发现,栽培黄瓜的YTH1起始密码子发生A→C突变,导致YTH结构域缺失,转录水平显著降低。通过基因敲除实验验证,YTH1是FL1.1的功能基因,且ACS2的栽培等位基因对YTH1存在隐性上位效应。这表明YTH1的有害突变通过影响m6A修饰的鉴定和调控,进而参与黄瓜果实长度的驯化过程。
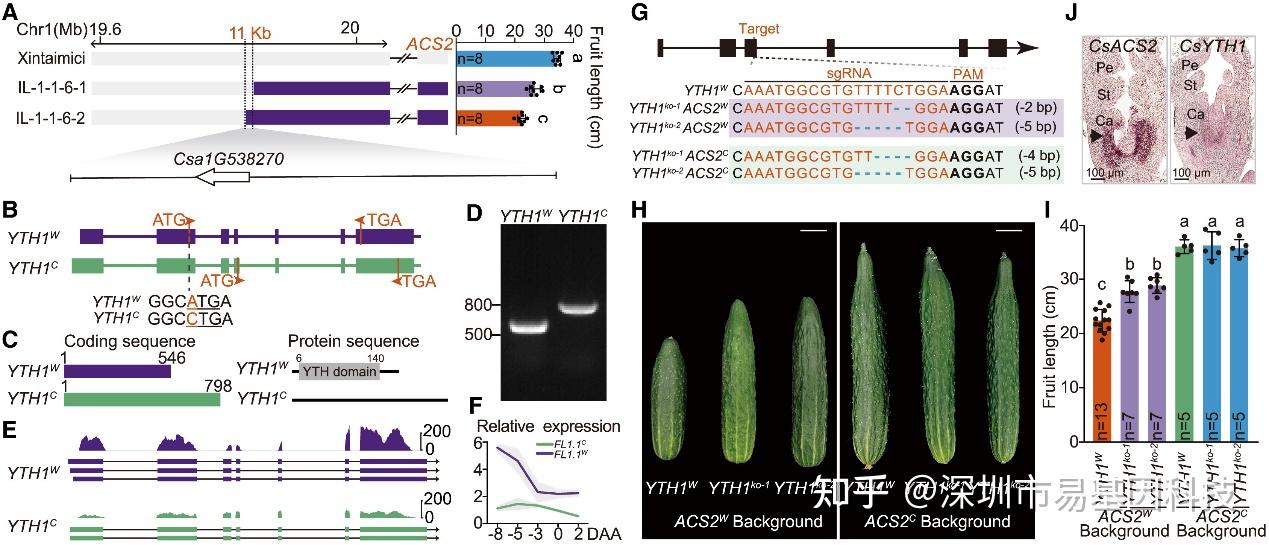
图3:FL1.1由YTH1基因的有害突变定义
- FL1.1的精细定位区间包含YTH1基因。上:选定重组体的基因型和果实长度。灰色和紫色分别代表Xintaimici和hardwickii的基因型。
- YTH1基因结构和变异。下方序列是起始密码子周围碱基。
- YTH1W和YTH1C的CDS和蛋白序列长度。数字表示CDS和YTH蛋白结构域的起始和终止位置。
- PCR显示YTH1W和YTH1C的CDS长度。
(E-F) 全长转录组(E)和RT-qPCR(F)显示mRNA的剪切和表达情况。
(G) 使用单RNA(single-guide RNA, sgRNA)通过CRISPR-Cas9技术生成YTH1ko突变体。sgRNA靶点和PAM分别用红色和加粗字体表示。缺失部分用蓝色虚线表示。
(H-I) YTH1ko突变体的果实长度。
(J) 原位杂交实验显示发育中的子房心皮中ACS2和YTH1 mRNA的表达情况。Pe表示花瓣;St表示雄蕊;Ca表示心皮。
(4)1287位C>T同义突变调控ACS2 mRNA m6A甲基化和蛋白翻译效率
通过m6A-seq分析发现,ACS2 mRNA的第三外显子存在显著的m6A peaks,且1287位C>T同义突变显著降低了ACS2 mRNA的m6A甲基化水平。进一步的免疫印迹和核糖体图谱分析显示,1287位C>T同义突变降低了ACS2蛋白水平和翻译效率(TE),而YTH1的存在进一步增强了ACS2¹²⁸⁷C的TE。这表明1287位C>T同义突变通过影响m6A修饰和YTH1的结合,调控ACS2的蛋白翻译效率,进而影响黄瓜果实长度。
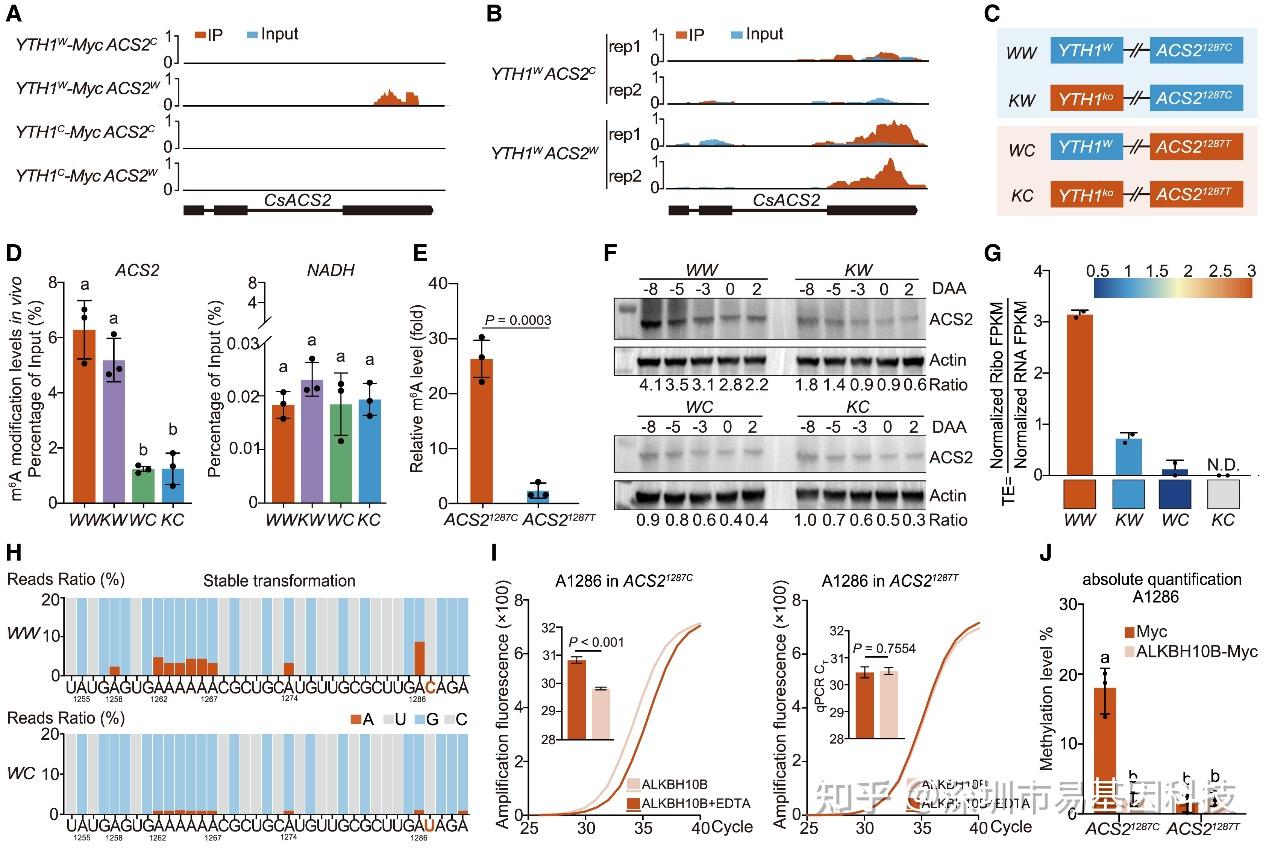
图4:1287位C>T 同义突变调控ACS2 mRNA m6A甲基化和蛋白TE
- ACS2 mRNA的FA-CLIP富集peak覆盖图。
- ACS2 mRNA的m6A-seq富集peak覆盖图。
- 四个等基因系的培育,仅在YTH1基因和ACS2 1287同义突变上存在差异。
- m6A-RIP-qPCR检测等基因系WW、KW、WC 和 KC中ACS2(左侧)和NADH(右侧对照)的相对m6A水平。
- m6A-RIP-qPCR 检测35S::ACS21287C 和35S::ACS21287T 的相对 m6A 水平。
- 四个等基因系中ACS2蛋白水平的免疫印迹分析。
- 核糖体分析法检测WW、KW、WC 和KC中ACS2的相对翻译效率(TE)。下:相应TE热图。
- 单碱基分辨率检测 ACS2 mRNA 1287同义SNP附近的 m6A 修饰。
- ACS21287C (左)和 ACS21287T(右)中A1286的m6A修饰SELECT检测结果。
- 绝对定量RT-PCR结果显示ACS21287C 中A1286的m6A修饰水平。
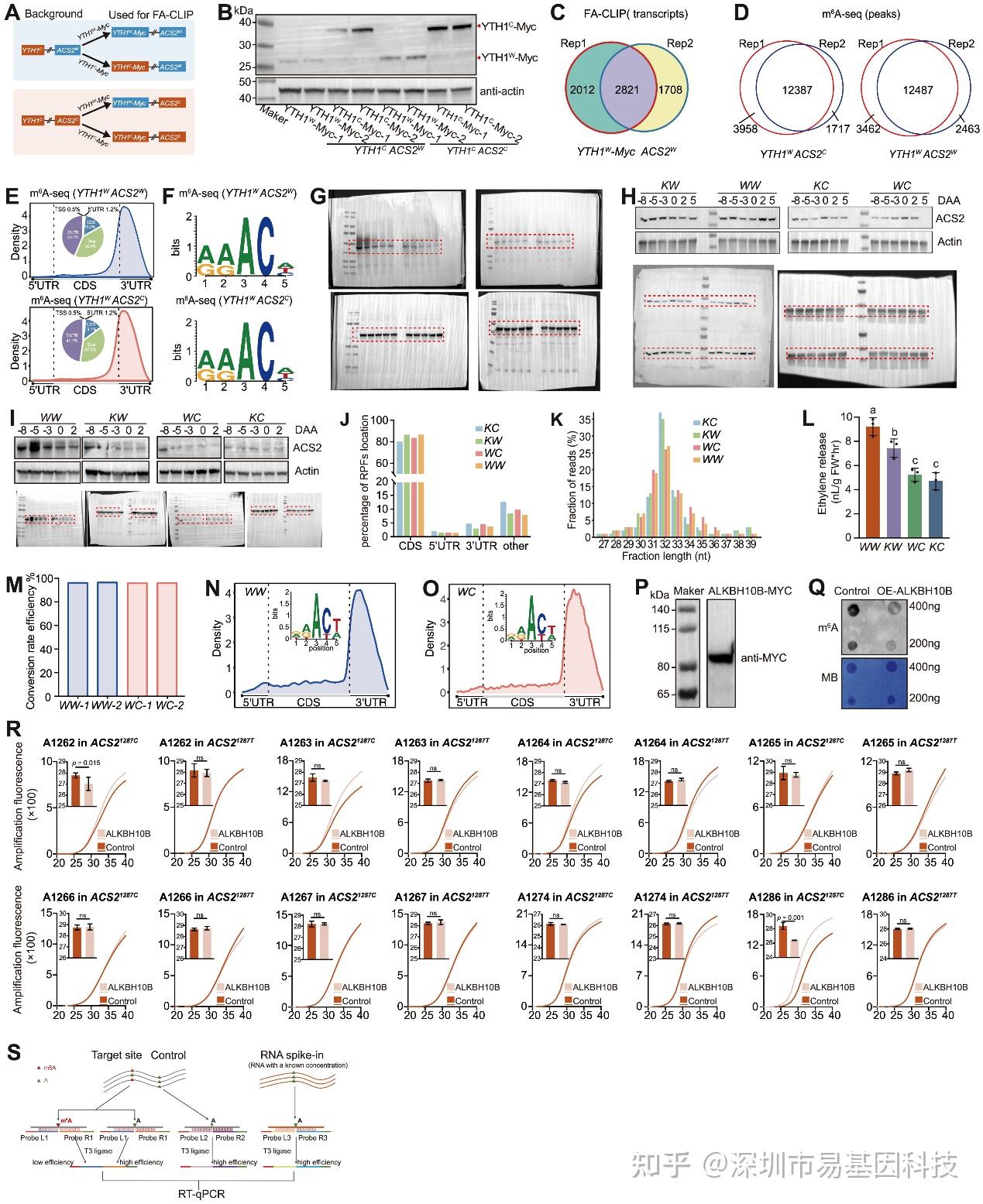
图S4:ACS2 mRNA中A1286的m6A甲基化检测,与图4相关
- FA-CLIP实验材料制备。
- 抗Myc抗体通过免疫印迹分析检测转基因材料中YTH1W-Myc 和YTH1C-Myc的蛋白表达。
- YTH1W-Myc W两个生物学重复的FA-CLIP中YTH1结合转录本重叠。
- YTH1W-Myc ACS2W和YTH1W-Myc ACS2C两个生物学重复m6A-seq中m6A甲基化peak重叠。
- m6A-seq鉴定的m6A位点在YTH1W-Myc ACS2W和YTH1W-Myc ACS2C转录本分布的meta基因分析。转录本分为三个区域:5′UTR、CDS和3′UTR。饼图显示在5′UTR、CDS和3′UTR区域分布的m6A位点比例(%)。
- 含有m6A的peak区域的保守序列motif。
(G–I) 与图4F相关的三个重复的未裁剪免疫印迹图像。
(J) 四种材料中核糖体足迹(RPFs)在CDS、5′UTR、3′UTR和其他区域的位置比例。
(K) RPFs长度比例分布。
(L) 四种基因型(WW、KW、WC和KC)中的乙烯产生量。
(M) 两个独立重复的eTAM-seq在转录组范围内的A→G转换效率。
(N-O) 通过eTAM-seq鉴定的m6A位点在WW和WC转录本中的分布的meta基因分析。
(P) 抗MYC抗体的免疫印迹分析显示ALKBH10B-MYC的表达。
(Q) 铜瓜同源的m6A去甲基化酶基因ALKBH10B过表达可以显著降低黄瓜子叶中的全局m6A水平。
(R) 检测ACS21287C和ACS21287T中1287同义突变周围的全部腺苷m6A修饰的SELECT结果。
(5)1287位C>T同义突变通过m6A修饰和RNA结构改变调控ACS2的TE
研究发现1287位C>T同义突变位于类似m6A保守motif DRACH的序列中,该突变破坏了m6A修饰位点,导致mRNA结构从松散状态变为紧凑状态,降低了YTH1与ACS2 mRNA的结合能力,最终使ACS2的TE下降。而野生型1287C通过m6A修饰形成松散RNA结构,结合YTH1后进一步偏向最弱构象,提高TE。这说明1287位C>T同义突变通过改变m6A修饰和RNA结构构象,精细调控ACS2翻译效率,从而决定黄瓜果实长度的驯化性状。
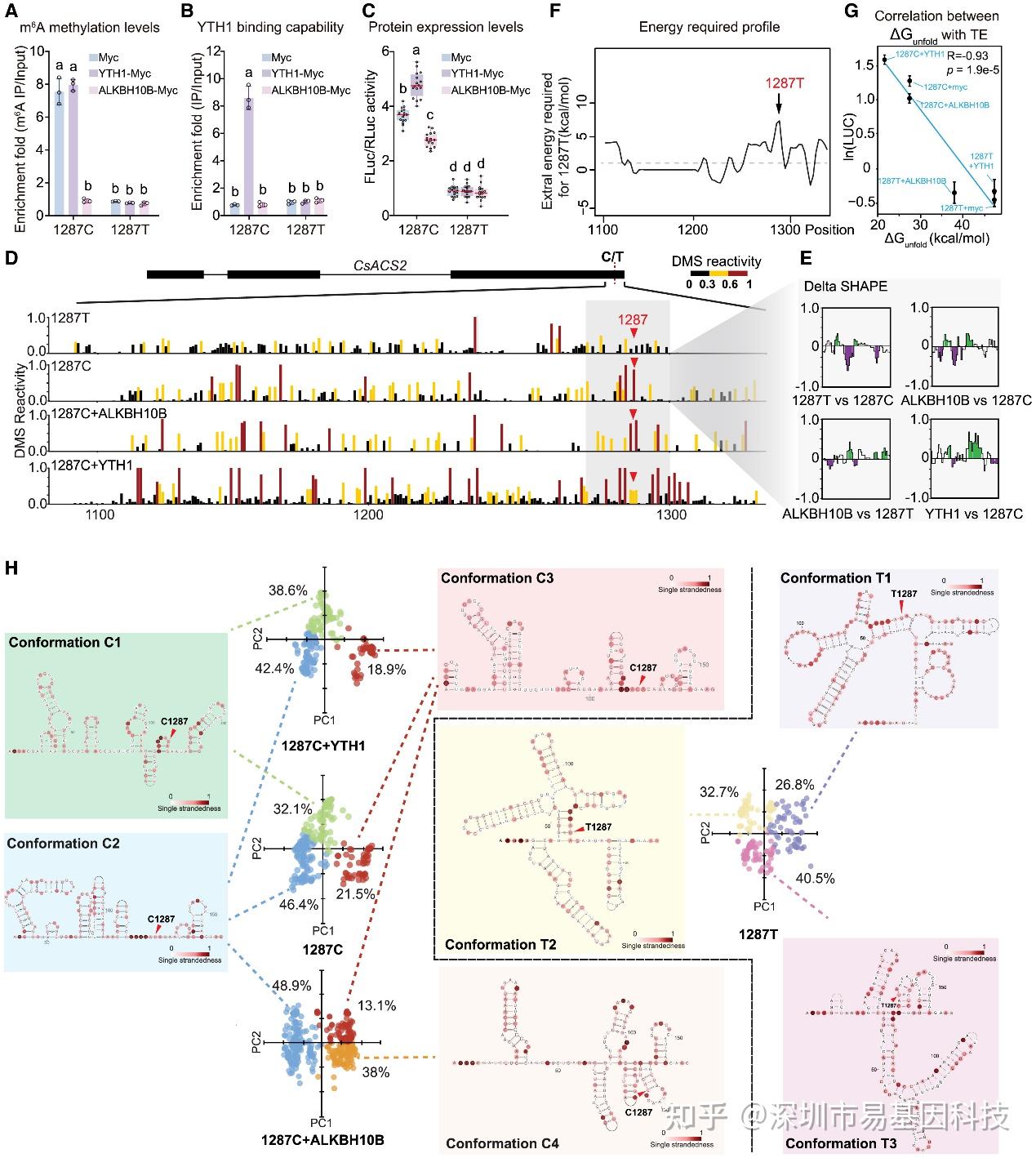
图5:1287位C>T同义突变通过m6A修饰和RNA结构改变调控ACS2的TE
- m6A-RIP-qPCR显示了与Myc、YTH1-Myc和ALKBH10B-Myc分别共表达时ACS21287C和ACS21287T的m6A甲基化水平。
- FA-CLIP显示了YTH1W与Myc、YTH1-Myc和ALKBH10B-Myc分别共表达时对ACS21287C和ACS21287T的结合能力。
- 双荧光素酶报告基因分析显示了与Myc、YTH1-Myc和ALKBH10B-Myc分别共表达时ACS21287C和ACS21287T的蛋白水平。
- ACS2等位基因的DMS反应性图谱。
- DeltaSHAPE图谱显示了(D)中灰色阴影区域的DMS反应性差异,计算为每种条件与后续条件之间的差异:ACS21287T与ACS21287C(1287T与1287C)、与ALKBH10B共表达的ACS21287C与ACS21287C(ALKBH10B与1287C)、与YTH1共表达的ACS21287C与ACS21287C(YTH1与1287C)以及与ALKBH10B共表达的ACS21287C与ACS21287T(ALKBH10B与1287T)。
- ACS21287T和ACS21287C等位基因之间打开RNA结构所需的能量差异。
- (C)中的TE与其对应的ΔG_unfold之间的皮尔逊相关性。
- CS21287T、ACS21287C以及分别与ALKBH10B或YTH1共表达的ACS21287C的代表性结构模型。结构通过主成分分析(PCA)可视化。
结论和启示
本研究通过遗传定位、基因克隆、基因编辑、m6A-seq分析等多种方法,揭示了黄瓜果实长度驯化性状的遗传调控机制,即ACS2基因中的1287位C>T同义突变通过改变m6A修饰和mRNA结构构象,调控ACS2蛋白的翻译效率,进而影响黄瓜果实长度。另外,YTH1基因的有害突变通过影响m6A修饰的识别和调控,也参与了果实长度的驯化过程。这一研究结果不仅挑战了同义突变无功能的传统认知,拓展了m6A在同义突变的研究领域,还为作物遗传改良提供了新的策略和思路。未来的研究可以进一步深入探索同义突变在其他生物性状调控中的作用,以及m6A修饰与其他RNA修饰在基因表达调控中的相互作用机制,为生命科学研究和农业生产实践提供更多的理论支持和技术指导。
m6A-seq技术的关键作用
本研究通过m6A-seq分析,精确地定位ACS2 mRNA上的m6A修饰位点,并发现1287位C>T同义突变显著降低了m6A修饰水平。这一发现为后续研究m6A修饰与RNA结构、蛋白翻译效率之间的关系提供了重要线索,进而揭示了m6A修饰在调控黄瓜果实长度驯化性状中的关键作用。
在今后类似的研究中,m6A-seq技术有望得到更广泛的应用。例如,在研究基因表达调控网络时,m6A-seq可以与其他组学技术(如转录组测序、蛋白质组测序等)相结合,全面解析m6A修饰在基因转录、剪接、翻译等过程中的作用,以及其与其他调控因子之间的相互作用关系。此外,m6A-seq技术还可应用于研究发育生物学、疾病发生机制等领域,通过分析不同发育阶段或疾病状态下m6A修饰的动态变化,寻找潜在的调控靶点和生物标志物,为疾病的诊断和治疗提供新的策略。
关于易基因RNA m6A甲基化测序(MeRIP-seq)技术
易基因MeRIP-seq技术利用m6A特异性抗体富集发生m6A修饰的RNA片段(包括mRNA、lncRNA等rRNA去除所有RNA),结合高通量测序,可以对RNA上的m6A修饰进行定位与定量,总RNA起始量可降低至10μg,最低仅需1μg总RNA。广泛应用于组织发育、干细胞自我更新和分化、热休克或DNA损伤应答、癌症发生与发展、药物应答等研究领域;可应用于动物、植物、细胞及组织的m6A检测。
大样本量m6A-QTL性状关联分析,传统MeRIP单个样品价格高,通常难以承担。易基因开发建立MeRIP-seq2技术,显著提成IP平行性,实现不同样本间相对定量,降低检测成本。
易基因提供适用于不同科研需求的MeRIP技术:
- m6A甲基化-常量mRNA 甲基化测序(MeRIP-seq)
- m6A甲基化-常量mRNA +lncRNA甲基化测序(lnc-MeRIP-seq)
- m6A甲基化-微量mRNA +lncRNA甲基化测序(Micro-lnc-MeRIP-seq)
- 高通量m6A甲基化-常量mRNA甲基化测序(MeRIP-seq2)
技术优势:
- 起始量低:样本起始量可降低至10-20μg,最低仅需1μg总RNA;
- 转录组范围内:可以同时检测mRNA和lncRNA;
- 样本要求:可用于动物、植物、细胞及组织的m6A检测;
- 重复性高:IP富集重复性高,最大化降低抗体富集偏差;
- 应用范围广:广泛应用于组织发育、干细胞自我更新和分化、热休克或DNA损伤应答、癌症的发生与发展、药物应答等研究领域。
研究方向:
m6A甲基化目前主要运用在分子机制的理论性研究
- 疾病发生发展:肿瘤、代谢疾病(如肥胖/糖尿病)、神经和精神疾病(如阿尔兹海默症/抑郁症)、炎症…
- 发育和分化:早期胚胎发育、个体/组织/器官生长发育、干细胞分化与命运决定、衰老
- 环境暴露与响应:污染、抗逆、生活方式
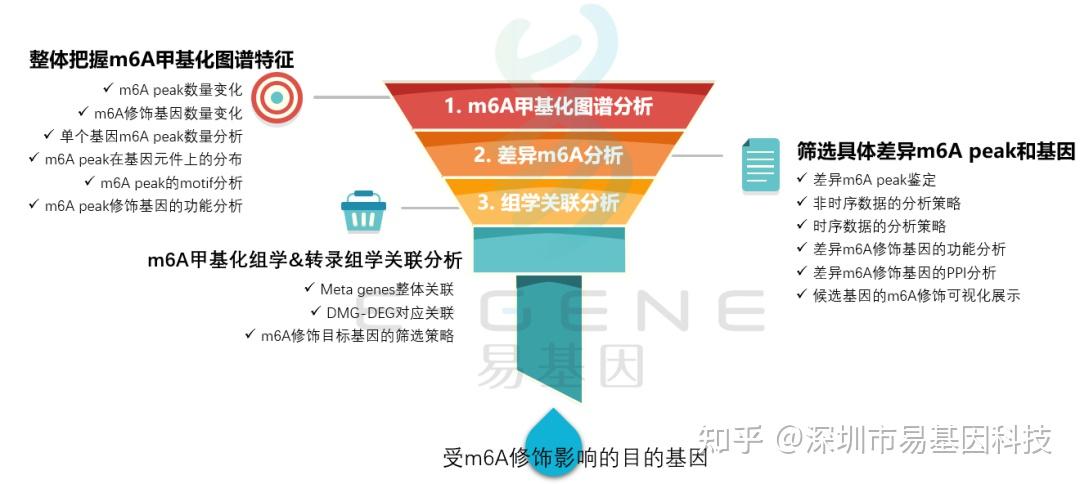
关于m6A甲基化研究思路
(1)整体把握m6A甲基化图谱特征:m6A peak数量变化、m6A修饰基因数量变化、单个基因m6A peak数量分析、m6A peak在基因元件上的分布、m6A peak的motif分析、m6A peak修饰基因的功能分析
(2)筛选具体差异m6A peak和基因:差异m6A peak鉴定、非时序数据的分析策略、时序数据的分析策略、差异m6A修饰基因的功能分析、差异m6A修饰基因的PPI分析、候选基因的m6A修饰可视化展示
(3)m6A甲基化组学&转录组学关联分析:Meta genes整体关联、DMG-DEG对应关联、m6A修饰目标基因的筛选策略
(4)进一步验证或后期试验
参考文献:
Xin T, Zhang Z, Zhang Y, Li X, Wang S, Wang G, Li H, Wang B, Zhang M, Li W, Tian H, Zhang Z, Xiao YL, Tang W, He C, Ding Y, Huang S, Yang X. Recessive epistasis of a synonymous mutation confers cucumber domestication through epitranscriptomic regulation. Cell. 2025 Jul 15. doi: 10.1016/j.cell.2025.07.006.
相关阅读:
1、2024项目文章精选:DNA甲基化、RNA甲基化(m6A/m5C)、ChIP-seq、单细胞转录组、宏基因组|年终盘点
2、项目文章 | Adv Sci:NSUN2介导m5C修饰代谢重编程促进肿瘤进展 揭示治疗新选择
3、项目文章 | NAR:RCMS编辑系统在特定细胞RNA位点的靶向m5C甲基化和去甲基化研究
4、项目文章:MeRIP-seq+RNA-seq揭示不同品种猪肌肉发育的m6A RNA甲基化差异|育种研究
5、项目文章:MeRIP-seq+RNA-seq揭示家禽(鸡)脂肪沉积中的m6A RNA甲基化调控机制
6、项目文章 | MeRIP-seq揭示m6A修饰在肺动脉高压(PAH)发病机制中的潜在作用和新治疗靶点
7、项目文章|EP/IF7.3:陆军军医大学刘晋祎团队揭示PM2.5暴露通过m6A甲基化调控雄性生殖功能损伤机制
8、项目文章|Hepatology/IF15.8:复旦中山医院沈英皓利用ChIP-seq及多组学解析肝癌仑伐替尼耐药机制(国人佳作)
9、项目文章 | 90天见刊,易基因m6A RNA甲基化(MeRIP)+转录组组学研究
10、项目文章|IJBM:安医大陈飞虎团队揭示METTL3介导m6A甲基化在炎症性疾病发病机制中的表观调控作用
11、项目集锦 | 易基因近期m6A甲基化(MeRIP-seq)研究成果
12、技术推介|RNA m6A甲基化测序(MeRIP-seq)技术介绍
农业生态专题